{kind=link}
{kind=link}
{kind=link}
{kind=link}
Last Chapter
Next Chapter
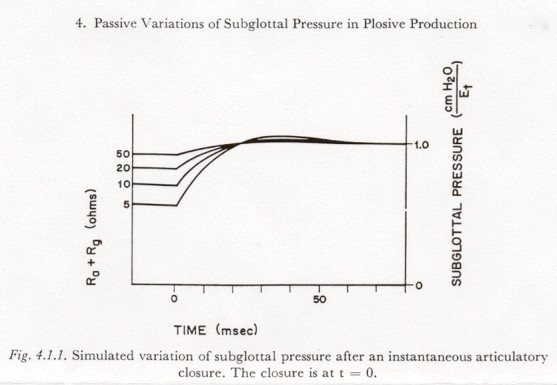
4. Passive Variations of Subglottal Pressure in Plosive Production
4.1 Subglottal Pressure Variations Due to an Articulatory Closing Gesture
In Section 3.1 above, the “passive” variations of subglottal pressure in a simple-released-plosive were defined as those caused by the variation of glottal-supraglottal air flow resistance during an articulatory closing gesture or release, or by a supraglottal cavity volume change. This section will discuss the variations in subglottal pressure that can be caused by a rapid articulatory closure.
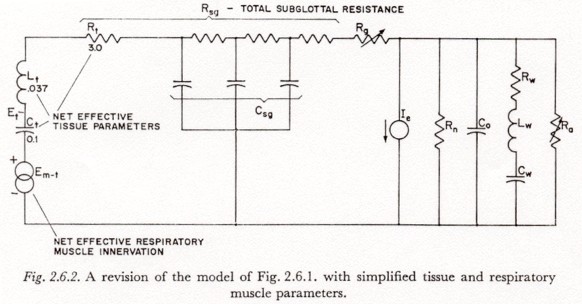
The variation in subglottal pressure after a rapid articulatory closure, with a constant respiratory muscle innervation, can be approximated by a family of curves of similar shape but varying amplitude, the amplitude being determined by the level of muscle innervation and the pre-closure glottal-supraglottal resistance. Figure 4.1.1 shows the variation in subglottal pressure in the model of Figure 2.6.2, after an essentially instantaneous opening of the circuit in the branch containing Ra, for a constant respiratory muscle innervation Em-t, and for a number of values of (Rg + Ra).
The post-closure portions of Figure 4.1.1 were obtained by simulation of the circuit of Figure 2.6.2, with Et held constant by a voltage source. (After closure, holding Et constant is essentially equivalent to holding Em-t constant.) The amplitude of the asymptotic total increase is equal to EtRsg / (Ra + Rg + Rsg) where Rsg is the total subglottal resistance.
The small pre-closure decay in pressure is due to the assumption of a constant muscle innervation (Em-t constant). The slope at closure is determined by the rate of discharge of Ct through (Rsg + Ra + Rg), and is equal to -Et / [Ct(Rsg + Ra + Rq)]
The preclosure pressure variations shown in Figure 4.1.1 are line segments drawn in at that slope. Figure 4.1.1 shows that the subglottal pressure rise after a fast (instantaneous) closure would have a rise time of about 20 msec with little, if any, overshoot. This time constant represents a lower limit for the time constants to be found in speech, and would only be attained for a change in air flow resistance that occurred in less than about 10 msec. As will be discussed in Chapter 5 below, the time constant of resistance change in a rapid articulatory closure can be less than 10 msec, but is often likely to be longer, depending on the place of articulation, phonetic environment, rate of speech, etc. For resistance-change time constants of more than about 40 msec (during an articulatory closing gesture), the subglottal pressure change will follow the change in articulatory resistance (with a small time delay). For resistance-change time constants between 10 and 40 msec (the range apparently most likely), however, the pressure variation will be a function of both the resistance change and the dynamic properties of the respiratory mechanism.
It is beyond the scope of this study to investigate experimentally the above theoretical observations concerning passive pressure variations. It has also been difficult to find a subglottal pressure record in the literature in which the passive pressure variation accompanying a closure for a plosive is clearly evident; this would be most likely in a sequence of a fricative or sibilant plus a plosive.
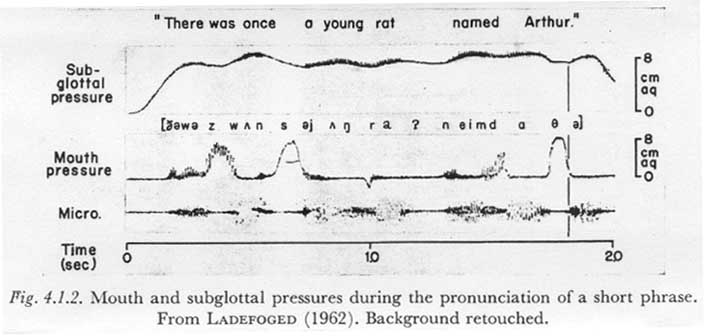
However, a related phenomenon occurred in the record shown in Figure 4.1.2, taken from a paper by LADEFOGED (1962).
LADEFOGED simultaneously recorded the subglottal pressure (indirectly via an esophageal balloon at the bifurcation of the trachea), the supraglottal (mouth) pressure, and the acoustic signal for the short phrase “There was once a young rat named Arthur”. From the evidence in these records, and a consideration of the possible actions of the speech mechanism, it seems safe to assume that at approximately the point in time indicated by the vertical broken line, a relatively heavy flow of air for the [q] was interrupted by a fairly rapid glottal closure marking the resumption of phonation. The air flow after this time (i.e., during the phonation), though not zero as during the closure of a plosive, would be relatively small.
The resulting perturbation in the subglottal pressure waveform is qualitatively clear, though the purely passive component of the variation is obscured by a changing respiratory innervation; before the closure, the subglottal pressure is fairly constant, indicating that it is being maintained by a rising net respiratory muscle innervation. This rising innervation is apparently no longer present at 50-75 msec after the closure. Compare the [q] with the [s] in “once”, for example, during which the respiratory innervation is probably fairly constant, resulting in a pressure decay due to the high air flow. (Rough calculations indicate that the rate of decay is in the range expected for the air flow of an [s] and a tissue compliance Ct of about 0.04.)
In order to obtain a quantitative relation between experimentally observed passive pressure variations and rapid changes of articulatory resistance, at least one more physiological variable is required in addition to the subglottal and supraglottal pressures; that is, a simultaneous recording of the volume velocity of the air flow is also needed. Further, it would be desirable to have instrumentation with a response time of less than 5 or 10 msec. A survey of the literature has revealed only one attempt to obtain such recordings (LADEFOGED, 1963), and some of the results of this experiment will be discussed in the next section.
4.2 Subglottal Pressure Variations Due to an Articulatory Release
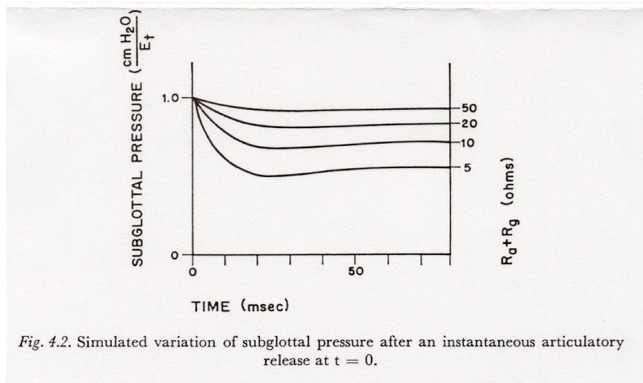
The study of the variation in subglottal pressure after the articulatory release of a simple-released-plosive is complicated by the fact that when the articulators have stopped opening, the glottal resistance might be in a period of rapid transition, i.e., the sum of (Ra + Rg) must in general be considered as having two simultaneously varying components. However, it is still instructive to neglect initially the fact that there are two components, and to look at the subglottal pressure response to a single fast decrease in the sum (Ra + Rg).
Figure 4.2 shows simulated variations in subglottal pressure for an instantaneous (step) change in (Ra + Rg) from � (open circuit) to the value shown as the parameter for each curve. The curves were recorded as the response to the closing of a switch in series with Ra in a simulation of the model of Figure 2.6.2. The net tissue tension Et, not the muscle innervation, was held constant with a voltage source. (Holding the muscle activation Em-t constant, instead of Et, mainly adds a decay to the output that tends to obscure the nature of the transients.)
As in the case of the response to a closure, the response-time is about 15-20 msec, and the asymptotic pressure change is EtRsg / (Ra + Rg + Rsg). However, in the case of a release, the shunting effect of (Ra + Rg) tends to reduce the response-time and increase the overshoot, especially for the smaller air flow resistances such as might be present after the release of an aspirated, unvoiced plosive. (This observation concerning overshoot vs. flow resistance may be compared with the observations of AGOSTONI and FENN mentioned below in this section.)
The time constant of the change in articulatory resistance might be either somewhat shorter or somewhat longer than the subglottal system response time of 15-20 msec. For example, the time constant may be shorter than 15 msec in the initial explosion of a closure made with the lips or tongue tip, or it may be somewhat longer in the post-explosion opening phase or in the release of a closure made with the blade or body of the tongue. These topics are discussed in more detail in Chapter 5.
There are a number of examples in the literature of the perturbation in subglottal pressure caused by the release of a plosive (for example, STETSON, 1951, Figures 19 and 20). As in the case of a perturbation caused by a closure, it is difficult to find studies in which a sufficient number of variables were recorded simultaneously, and with sufficient accuracy, to enable a quantitative evaluation of the parameters of an underlying model of the respiratory system. However, one such study was reported by LADEFOGED (1963). During the production of short English sentences by a number of speakers, LADEFOGED recorded simultaneously the subglottal pressure (using the esophageal technique mentioned previously), the supraglottal pressure, and the air flow (using a technique in which the deflection of a rubber diaphragm in the outlet of a mask is measured optically). He reports a drop in subglottal pressure of about 10 % during the release of a voiceless, aspirated stop. In one record of this nature shown in the article, a drop in subglottal pressure of about one to one and one-half cm H2O was accompanied by an average air flow of about one 1/sec. These data indicate that the effective total subglottal resistance is in the order of one to one and one-half Ohms. This figure is at variance with the estimates of about four Ohms assumed in this paper and by VAN DEN BERG, and indicated in a number of places in the respiration literature. The cause of the discrepancy is not clear and further work in this area is necessary.
To circumvent the difficulty of making numerous simultaneous physiological measurements, the passive variation of subglottal pressure with changes of glottal-supraglottal resistance can be determined experimentally by using non-speechlike maneuvers in which certain variables are naturally relatively constant or are externally controlled. A common experimental methodology in the study of respiration is suddenly to open or close a valve in a line containing a fixed, calibrated air flow resistance, while the subject is expiring (or inspiring) into the line at a relatively constant pressure. If subglottal pressure is measured simultaneously, fairly direct comparisons can be made to the theoretical results in Figure 4.1.1 and 4.2.
However, in the past little attention has been paid to the transient response in such maneuvers, the interest being more in the values after the transient has decayed. A study of this nature has been reported by AGOSTONI and FENN (AGOSTONI and FENN, 1963). These authors have noted some “overshoot” in the alveolar 1 pressure transient resulting from opening a closed valve. This phenomenon is noted to be most predominant in “the tests without added flow resistances” (besides those of the valve and flow measuring equipment). However, since the transient was not of direct interest in their study, the report contains little beyond a mention of the phenomenon and a reference to the possibility that “proprioceptive reflexes” may be elicited at the release.
4.3 Subglottal Pressure Variations Due to a Supraglottal Cavity Volume Change
In the model of Figure 2.6.2 it can be seen that a current Ie (an 'active' change in volume of the supraglottal cavity) will cause a change in subglottal pressure under the conditions of a complete articulatory and velopharyngeal closure and a non-infinite Rg (glottis not tightly closed). In this section we will investigate the extent of the possible subglottal pressure variations under these conditions.
Some proportion of a supraglottal volume increase might be due to a movement which simultaneously shortens the trachea and therefore decreases the subglottal volume. However, in the following discussion this effect will be ignored.
Referring to the simplified model of Figure
2.6.2, we note that except for transient effects introduced by Co,
Cw, and Csg, the current Ie can be considered
to act on Rsg, Lt and Ct in series. The change
in subglottal pressure caused by Ie is given by: IeRsg
+ Lt dIe / dt + (1/Ct)ò
Ie dt
In Chapter 7, evidence will be presented to support the conjecture that the maximum increase in supraglottal cavity volume that may be expected during the period of closure of a voiced plosive is about 15 ml and that this change can occur within about 0.1 sec. If we also assume that the onset and offset times of the rate of change of volume' (Ie) are both about 35 msec, the following three observations can be made :
(1) The onset and offset times are sufficiently long that the inductive term can be neglected without much error. The change of subglottal pressure is then given by: IeRsg + (1/Ct) ò Ie dt
(2) The term IeRsg can be as large as 0.15 X 3 = 0.45 cm H2O (assuming an average flow of 0.015 liter in 0.1 sec, or 0.15 liter/sec).
(3) The term (1/Ct) ò Ie dt can be as large as (1/0.1) (0.015) = 0.15 cm H2O, assuming Ct = 0.1. If Ct is lower, say equal to 0.05, this term can be as much as 0.3 cm H2O. This component will gradually increase during the duration of a current pulse and remain after the termination of Ie (or the release of the plosive, whichever comes sooner).
The maximum change in subglottal pressure that may be expected from an active increase in supraglottal cavity volume during a period of articulatory and velopharyngeal closure is then about 0.75 cm H2O. When the volume change is not extreme, it may be expected that the maximum change in subglottal pressure will be less than one-half cm H2O. Changes of this magnitude are probably not of much significance in speech but might be occasionally visible on a record of subglottal pressure.
Last Chapter |
Next Chapter |